The Bacterial Cell Interior.
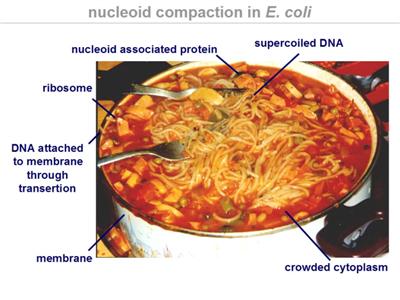
Chapter 3 of Microbe explains how the cell interior of bacteria is a very busy, crowded, and viscous place. A major component taking up space in this region is the nucleoid, and there is much more that can be said about it than there is space for in the chapter.
The DNA of the nucleoid is heavily folded and super-coiled. The DNA of Escherichia coli, if it were uncoiled, would be about 1000 times the length of the cell. Bacteria are thus very compact with genetic information storage.
An interesting point made by Schaechter in Chapter 3 that transcription of DNA into RNA only occurs at the nucleoid-cytosol interface, so organisation of the nucleoid's coiled domains is functionally important. Diagrams of this are presented in teaching sessions.
Recently, many dramatic advances have been made in scientific understanding of bacterial cytoplasmic filament proteins such as MreB and FtsZ , and these are already mentioned in chapter 2 of Schaechter.
Study Question:
How were MreB and FtsZ proteins first identified, and where do their names come from? Where are they located in the cell?
posted by GMO Pundit at 11:06 am
![]()
![]()

10 Comments:
Regarding its location;
MreB filaments is right beneath the bacterial cell membrane and give bacteria a strong, rigid structure and the rod shape of many bacteria.
FtsZ forms the Z-ring that determines the septum of bacterial cell division. Usually assembled in the center of the bacteria.
This is good research for answers.
I would question that MreB itself gives bacteria rigidity. The murein sacculus does that. MreB does in some way determine rod-like shape rather than a coccus shape.
Both names come about because they were names for classes of MUTANT bacteria of E. coli. Mutations in functions are a very useful tool for finding out details of unknown functions inside cells.
Mutations with a PHENOTYPE affecting physical shape are likely to be in genes connected with the cell wall or its formation pathways.
NEXT question:
Both types of work stated with Temperature sensitive mutants.
What does temperature-sensitive mean, and how are temperature sensitive mutants useful?
MICROBE PUNDIT
archeak9 has missed one ESSENTIAL point I'm looking for about ts mutants, so there is more work for other contributors still.
THIRD STUDY QUESTION
How do we know that Staphylococcus doesn't have a MreB homologue protein?
In a recent study (Jones et al. ), they used immunofluorescence to visualize elongated polymers of MreB and closely related protein Mbl that encircle the cell as large spirals under the cell membrane. They are widely distributed among rod-shaped, filamentous, and helical bacteria, suggesting that an MreB cytoskeleton is important to generate a non-spherical shape.As we know , staphylococci are cocci (spehrical shape) thus they do not have Mre B and its homologue protein.
Fam that is a prediction about Staph not an observation.
My point is that the complete gene sequence of Staph enables us to know that mreB homologue genes are absent, because we can directly interogate the sequence with known mreB genes from say G+ve Bacillus, using BLASTP software for instance.
Note the nomemclature for genes is different for proteins MreB.CAPITALISED NON ITALIC
Good work Fam, putting in with answers.
how are temperature sensitive mutants useful?
ts mutants are useful to find out the essential function that important for its survival. We can find mutants that are alive but defective in survival function by adding the stuff they need in growth medium.
Fam:
For an exam question it is important to demonstrate that you understant the issue invoved. From your answer is is very difficult to work out if you understand why ts mutants are so useful.
The most condusing part of your answer is where you say "We can find mutants that are alive but defective in survival function by adding the stuff they need in growth medium".
The context in which you put this sentance makes me think you don't undestant ts mutants very well.
This is an important point.
Lets take the ts mutant that allowed the gene ftsZ to be found in the first place.
My question:
Will the ftsZts mutant's growth ever be restored at the non-permisive temperature (37 deg C) BY ADDING FtsZ protein to the growth medium?
If so explain to us how FtsZ PROTEIN gets inside the cell where it is needed.
no, we can't restore back the ftsZts mutant by adding the protein because they won't get into the cells. I do know that for some structure defectiveness , we can't add something in the medium to let them grow. But i'm a bit confused because during your lecture on cell cycle , u said if we want to find out mutants that are alive but defective in survival function, we can do so by adding the stuff they need in growth medium. Is that correct?
However, i've realised i've put an irrelevant answer on usefulness of ts mutant. I'm sorry for that. I'm now trying to answer for second time.If we have a temperature-sensitive mutation in the gene of interest. We can have them grow normally in permissive temperature and by disable the gene product at high temperature , to discover the gene function. Let's take FstZ Ts mutant as an example, we observe them during high temperature, forming long filaments and comparing their normal phenotype during permissive temperature, we can know the function of FstZ protein is involved in cell division.
Perhaps I was not clear enough in my talk. Ill explain it further , hopefully better.
There are some types of mutant defect that can be corrected by adding extra nutrients to the growth medium. An example is a defect in histidine biosynthesis (his -ve mutants), because this can be corrected by adding histidine to the growth medium. This is successful because cells have natural mechanisms to take up histidine from the growth medium.
However the cels do not have mechanisms to take up proteins like FtsZ and MreB from the growth medium.
To help find mutants affecting these essential functions ts mutations are very useful, as they are not defective at low temperatures, and can be grown successfully at lower temperatures.
Good work Fam.
Post a Comment
<< Home