Quorum Sensing in the pathogenicity of Pseudomonas aeruginosa
M. I-S Low
Abstract
Pseudomonas aeruginosa is a Gram negative, motile, anaerobic rod that belongs to the genus pseudomonads. It is known for its versatility for survival in a range of ecological niches. This bacterium inhabits soil, water and vegetation. It is an opportunistic pathogen in humans, causing a variety of diseases such as urinary and gastrointestinal tract infections, and respiratory system infections. In immunocompromised patients infected with HIV or cystic fibrosis (CF), this pathogen is found to cause a mortality rate of about 50% (Todar, 2004). The bacteria can be isolated from the skin, throat and stool. It is spread by fomites and contaminated water (Arevalo-Ferro et al, 2003). P. aeruginosa is known for its resistance to many antibiotics because of the presence of the lipopolysaccharide, which prevents host immune cells from recognising it. In addition, the ability to form biofilms by the bacteria makes the cells more resistant to antibiotics as higher concentrations are needed to disrupt biofilm formation (Todar, 2004). P. aeruginosa has a large genome, consisting of over 6 million base pairs and over 5000 ORFs, encoding cellular genes (Stover et al., 2000). The versatility of the organism is most probably attributed to its large genome size and complexity.
Quorum sensing (QS) is a mechanism whereby bacteria communicate with one another, relying on bacterial population density. Gram negative bacteria, such as P. aeruginosa rely on N-acyl-homoserine lactones (AHL) as signal molecules in QS systems (Rasmussen et al., 2006). The paradigm of quorum sensing states that AHLs are constitutively produced at low cell densities. The AHLs will then accumulate in the environment, in proportion to the increase in bacterial population. At a certain threshold concentration of AHL, the molecules will be able to bind to its respective receptor and a series of target gene regulations will be activated (Juhas et al., 2005). This system of regulation ensures that bacteria are able to form organised communities and to exchange information with other members to coordinate cellular activities. Among the processes regulated by quorum sensing are the synthesis of secondary metabolites, enzymes and virulence factors which allow bacteria to colonize various ecological niches (as reviewed by Juhas et al. 2005).
In P. aeruginosa, the quorum sensing systems are made up of the two AHL systems: las and rhl systems; and one non-AHL system: 2-heptyl-3-hydroxy-4-quinolone (PQS) (Pesci et al, 1999). The QS systems in P. aeruginosa regulate about 6-10% of the bacterial genome, indicating that this system play an important role in the survival of the bacteria (Arevalo-Ferro et al., 2003, Todar, 2004). QS in P. aeruginosa is important in pathogenicity as it ensures the bacteria do not express pathogenic traits until population has reached its critical density to be able to overwhelm host defences and establish an infection (Arevalo-Ferro et al., 2003). Among the virulence factors regulated by QS are proteases, exotoxin A, rhamnolipids, pyocyanin and sideophores (Wagner et al., 2003). In this report, the QS system is discussed in terms of its importance in the virulence of the organism as it causes serious implications in immunocompromised patients.
Mode of synthesis
The las system in P. aeruginosa quorum sensing comprise of LasI and LasR, where LasI synthesises the signal molecule N-(3-oxo-dodecanoyl)-L-homoserine lactone (3-oxo-C12- HSL) and LasR acts as a transcriptional regulator.
The second quorum sensing system, the rhl system, is comprised of RhlI and RhlR, where RhlI synthesises the production of signal molecule N-butanoyl-L-homoserine lactone (C4-HSL) and RhlR is the transcriptional regulator (Erikson et al., 2002, Rasmussen et al., 2006). LasR and RhlR are able to induce transcription of their own genes, creating a positive feedback loop that increases AHL production and dissemination (Schuster et al., 2006).
The PQS signalling system links the las and rhl systems. LasI and RhlI have been shown to control the synthesis of PQS which in turn, controls the expression of RhlI and RhlR. PQS gene transcription is positively regulated by LasR, and at the same time, it is negatively regulated by rhl. Therefore, the ratio of 3-oxo-C12-HSL:C4-HSL is important (as reviewed by Juhas et al., 2005). The las and rhl systems do not act independently; they are arranged in a hierarchical fashion where 3-oxo-C12-HSL positively regulates the rhl system (Erikson et al., 2002). Figure 1 represents a simplified depiction of the las and rhl system.

Figure 1 Quorum sensing in P. aeruginosa. The las act in a hierarchical fashion, whereby it controls the rhl. The LasR/3-oxo-C12-HSL complex activates the transcription of rhlR. Additionally, 3-oxo-C12-HSL can also block activation of RhlR by C4-HSL. Both quorum signaling systems regulate the expression of various genes (lasB: LasB elastase, lasA: LasA elastase, toxA: exotoxin A, aprA: alkaline protease, xcpP and xcpR: genes of the xcp secretory pathway, rhlAB: rhamnosyltransferase and rpoS: stationary phase sigma factor (Van Delden et al., 1998).
Structure
The structures of the AHL molecules and the PQS molecule are shown in Figure 2.

Fig. 2. Structures of PQS and AHLs exploited for cell-to-cell communication by P. aeruginosa (Juhas et al., 2005).
Biological Roles
The las system is mainly involved in the regulation of various virulence factors, while the rhl system regulates a broad spectrum of P. aeruginosa genes (Juhas et al., 2005). In CF patients, lasR transcripts have been detected in the sputum samples, and this accumulation correlated with lasA, lasB, toxA, arpA, lasI and rhlR mRNA transcripts, which are virulence factors (Pesci et al, 1997, Rumbaugh et al, 1999), indicating a functional link on the regulation of these genes. RhlR has been shown to bind to a specific upstream sequence of the rhlAB gene independently of the presence or absence of C4-HSL. When C4-HSL is present, transcription of rhlAB is activated, while transcription is repressed in the absence of C4-HSL (as reviewed by Juhas et al., 2005). C4-HSL has also been shown to regulate virulence genes such as lasB, rpoS, rhlA and rhlI (Rumbaugh et al., 1999). The PQS signalling system is able to exert antimicrobial activity and is shown to promote biofilm formation (as reviewed by Juhas et al., 2005).
Quorum sensing also play a part in host-pathogen interactions by modulating the host immune system. The AHL mainly responsible in immunomodulation of the immune system is 3-oxo-C12-HSL (Wagner et al., 2006). It is able to induce inflammation in infected hosts, which produces further damage. When 3-oxo-C12-HSL is produced, immune cells, such as lymphocytes, macrophages and antibodies are activated. The long chain AHL of 3-oxo-C12-HSL suppresses IL12 and TNFα secretion, skewing the T cell response to a Th2 type response (Telford et al., 1998). 3-oxo-C12-HSL has also been shown to induce Cox2 [an inflammation associated enzyme in macrophages]. Cox2 induces inflammation, fever and pain, which enables the pathogen to disseminate and cause septicemia. Therefore, this shows that 3-oxo-C12-HSL does not only regulate virulence genes, but it is a virulence factor by itself (Smith et al., 2002). 3-oxo-C12-HSL, which controls PQS production through the regulation of pqsH show that these compounds synergise to reduce T cell proliferation at sub-cytotoxic doses (Pritchard, 2006). PQS is able to act in the T cell signalling pathway by enhancing IL2 production, which in turn inhibits T cell proliferation. This suggests that the pathogen is capable of influencing the immune system to optimise its survival (Pritchard 2006, Smith et al., 2003).
To show that 3-oxo-C12-HSL and C4-HSL are directly accountable for pathogenicity, mutations in the las and rhl systems were made and the effects on pathogenicity were observed. Various experiments have shown that there is a general decrease in virulence in these mutants. In lung infection models, a lasI and rhlI double mutant produced less virulence factors, resulting in a faster immune response, stronger oxidative bursts of blood PMNs and accumulate antibodies faster (Rasmussen et al., 2006). In burn wound infections, lasI and rhlI double mutants showed reduced virulence as the pathogen was less efficient in dissemination compared to its wild type counterpart. This is because virulence factors, such as alkaline protease and elastase which interfere with phagocytosis of neutrophils are absent (Rumbaugh et al., 1999). In acute pulmonary infections (Pearson et al., 2000), burn wound infections (Rumbaugh et al., 1999) and chronic lung infections (Wu et al., 2001), mutants in QS genes have been able to reduce mortality compared to wild type infections. In a study done by Arevalo-Ferro et al. (2003), lasI and rhlI double mutant showed that cellular protein composition was significantly reduced compared to the cell’s transcriptome. This suggests that the LasR and RhlR transcriptional regulators are able to activate or repress the transcription of target genes. However, it should be noted that double mutants are not completely avirulent. Virulence of the bacteria is multi-factorial and quorum sensing is only a system that plays a part in virulence (Smith et al., 2003).
Practical Applications
The understanding of quorum sensing in P aeruginosa is important as it controls one third of the virulence genes. Hence, it can be exploited as a target for antimicrobials. Mutations made in the QS regulatory genes have been shown to reduce virulence and mortality. There are three ways to interfere with the QS system: blocking AHL synthesis, degrading the AHL signal molecule or inhibiting the binding of the signal molecule to the receptor (Juhas et al., 2005, Rasmussen et al., 2006). These methods minimise the selective pressure for resistant bacteria as it does not affect its growth (Juhas et al., 2005).
As precursors such as acyl-ACP and SAM are used in the synthesis of the AHL molecules, analogues of SAM, such as Holo ACP, L/D-S-adenosyl-homocystein, sinefugin and butyryl-SAM can inhibit synthesis of AHL synthetases, such as RhlI (as reviewed by Juhas et al., 2005). Therefore, there will be no accumulation of AHLs despite the increase in bacterial population. The AHL signal molecule could be degraded using chemical, enzymic or metabolic methods (Rasmussen et al., 2006). AiiA enzyme from Bacillus sp can inactivate AHLs by hydrolysing the lactone bond, also known as quorum quenching (as reviewed by Juhas et al., 2005).
Lactonolysis, or ring opening of the AHL lactones could be triggered by high pH as well as high temperatures. These result in lowered amounts of bioactive AHLs (Lee et al., 2002). Activation of the transcription regulators could be blocked using analogues of signalling molecules. Substitutions at the 3-oxo-C6-HSL acyl side chain are able to displace C6-HSL from activating the regulator (as reviewed by Juhas et al. 2005). The HSL ring can also be replaced with an alternative ring structure, such as exchanging the lactone ring for the amino cycloalcohol or amino cycloketone. These were shown to inhibit QS controlled expression of a plasI-gfp fusion and selected virulence factors (Smith et al., 2003).
Antimicrobials produced by other organisms could also block QS signalling. Delisea pulchra produces furanone, which is able to block cell-to-cell communication and is shown to be able to clear P. aeruginosa lung infections in mouse models (as reviewed by Juhas et al., 2005). QS inhibitors, such as para-benzoquinone and 4-nitro-pyridine-N-oxide (4-NPO) can also be used to target RhlR and LasR (Rasmussen et al., 2005). Eukaryotic organisms, such as fungi have also been shown to produce antimicrobial compounds such as patulin and penicillic acid to downregulate 45-60% of QS regulated genes.
QS systems are important in regulating the virulence of P. aeruginosa. Therefore, continued research to produce novel antibiotics that will be able to efficiently target this system would be greatly beneficial to reduce nosocomial infections and reduce mortality in immunocompromised patients.
References
Arevalo-Ferro, C., M. Hentzer, G. Reil, A. Görg, S. Kjelleberg, M. GIvskov, K. Riedel, and L. Eberl. 2003. Identification of quorum-sensing regulated proteins in the opportunistic pathogen Pseudomonas aeruginosa by proteomics. Environmental Microbiology. 5: 1350-1369.
Erikson, D. L., R. Endersby, A. Kirkham, K. Stuber, D. D. Vollman, H. R. Rabin, I. Mitchell, and D. G. Storey. 2002. Pseudomonas aeruginosa quorum-sensing systems may control virulence factor expression in the lungs of patients with cyctic fibrosis. Infection and Immunity. 70: 1783-1790.
Heurlier, K., V. Dénervaud, and D. Haas. 2006. Impact of quorum sensing on fitness of Pseudomonas aeruginosa. International Journal of Medical Microbiology. 296: 93-102.
Juhas, M., L. Eberl, and B. Tümmler. 2005. Quorum sensing: the power of cooperation in the world of Pseudomonas. Environmental Microbiology. 7: 459-471.
Lee, S. J., S.Y. Park, J.J. Lee, D.Y. Yum, B.T. Koo and J.K. Lee. 2002. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Applied Environmental Microbiology. 68: 3919–3924.
Pearson, J. P., M. Feldman, B. H. Iglewski, and A. Prince. 2000. Pseudomonas aeruginosa cell-to-cell signalling is required for virulence in a model of acute pulmonary infection. Infection and Immunity. 68: 4331-4334.
Pesci E. C., Pearson J. P., Seed P. C., Iglewski B. H. 1997. Regulation of las and rhl quorum sensing in Pseudomonas aeruginosa. Journal of Bacteriology 179:3127–3132
Pritchard, D. I. 2006. Immune modulation by Pseudomonsa aeruginosa quorum-sensing signal molecules. International Journal of Medical Microbiology. 296: 111-116.
Rasmussen, T. B., and M. Givskov. 2006. Quorum-sensing inhibitors as anti-pathogenic drugs. International Journal of Medical Microbiology. 296: 149-161.
Rasmussen, T. B., T. Bjarnsholt, M.E. Skindersoe, M. Hentzer, P. Kristoffersen, M. Kote, J. Nielsen, L. Eberl and M. Givskov. 2005. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. Journal of Bacteriology. 187: 1799–1814.
Rumbaugh, K. P., J. A. Griswold, B. H. Iglewski, and A. N. Hamood. 1999. Contribution of quorum sensing to the virulence of Pseudomonas aeruginosa in burn wound infections. Infection and Immunity. 67: 5854-5862
Schuster, M., and E. P. Greenberg. 2006. A network of networks: quorum sensing gene regulation in Pseudomonas aeruginosa. International Journal of Medical Microbiology. 296: 73-81.
Smith, R. J., S. G. Harris, R. Phipps, and B. Iglewski. 2002. The Pseudomonas aeruginosa quorum-sensing molecule N-(3-Oxododecanoyl)Homoserine Lactone contributes to virulence and induces inflammation in vivo. Journal of Bacteriology. 184: 1132-1139.
Smith, R. S., and B. H. Iglewski. 2003. P. aeruginosa quorum-sensing systems and virulence. Current Opinion in Microbiology. 6: 56-60.
Stover, C. K., X. Q. Pham, A. L. Erwin, A. L. Erwin, S. D. Mizoguchi, P. Warrener, M. J. Hickey, F.S. L. Brinkman, W. O. Hufnagle, D. J. Kowalik, M. Lagrou, R. L. Garber, L. Goltry, E. Tolentino, S. Westbrock-Wadman, Y. Yuan, L. L. Brody, S. N. Coulter, K. R. Folger, A. Kas, K. Larbig, R. Lim, K. Smith, D. Spencer, G. K.-S. Wong, Z. Wu, I. T. Paulsenk, J. Reizer, M. H. Saier, R. E. W. Hancock, S. Lory, and M. V. Olson. 2000. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature. 406: 959-964
Telford, G., D. Wheeler, P. Williams, P.T. Tomkins, P. Appleby, H. Sewell, G. Stewart, B.W. Bycroft and D.I. Pritchard, 1998. The Pseudomonas aeruginosa quorum-sensing signal molecule N-(3-oxododecanoyl)-L-homoserine lactone has immunomodulatory activity. Infection and Immunity. 66: 36–42.
Todar’s Online Textbook of Bacteriology. http://textbookofbacteriology.net/pseudomonas.html
Van Delden, C., and B. H. Iglewski.1998.Cell-to-cell signaling and Pseudomonas aeruginosa infections. Emerging Infectious Diseases 4: 551-560
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I. & Iglewski, B. H. 2003. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. Journal of Bacteriology. 185: 2080–2095.
Wagner, V. E., J. G. Frelinger, R. K. Barth, and B. H. Iglewski. 2006. Quorum sensing: dynamic response of Pseudomonas aeruginosa to external signals. Trends in Microbiology 14: 55-58
Wu, H., Z. Song, M. Givskov, G. Doring, D. Worlitzsch, K. Mathee, J. Rygaard, and N. Høiby. 2001. Pseudomonas aeruginosa mutations in lasI and rhlI quorum sensing systems result in milder chronic lung infection. Microbiology. 147: 1105-1113.
Abstract
Pseudomonas aeruginosa is a bacterium responsible for a wide range of infections. Quorum sensing (QS) is used in P. aeruginosa as a means of cell-to-cell communication and is particularly important in its pathogenicity. The QS system is composed of two AHL systems: las and rhl; and a non-AHL system: PQS. The involvement of these systems in the virulence of this pathogen is discussed in this report. It has been observed that quorum sensing play an important part in the virulence of this pathogen. Therefore, it is important that novel antimicrobials should be able to target this system to reduce virulence in animal model infections.Introduction
Pseudomonas aeruginosa is a Gram negative, motile, anaerobic rod that belongs to the genus pseudomonads. It is known for its versatility for survival in a range of ecological niches. This bacterium inhabits soil, water and vegetation. It is an opportunistic pathogen in humans, causing a variety of diseases such as urinary and gastrointestinal tract infections, and respiratory system infections. In immunocompromised patients infected with HIV or cystic fibrosis (CF), this pathogen is found to cause a mortality rate of about 50% (Todar, 2004). The bacteria can be isolated from the skin, throat and stool. It is spread by fomites and contaminated water (Arevalo-Ferro et al, 2003). P. aeruginosa is known for its resistance to many antibiotics because of the presence of the lipopolysaccharide, which prevents host immune cells from recognising it. In addition, the ability to form biofilms by the bacteria makes the cells more resistant to antibiotics as higher concentrations are needed to disrupt biofilm formation (Todar, 2004). P. aeruginosa has a large genome, consisting of over 6 million base pairs and over 5000 ORFs, encoding cellular genes (Stover et al., 2000). The versatility of the organism is most probably attributed to its large genome size and complexity.
Quorum sensing (QS) is a mechanism whereby bacteria communicate with one another, relying on bacterial population density. Gram negative bacteria, such as P. aeruginosa rely on N-acyl-homoserine lactones (AHL) as signal molecules in QS systems (Rasmussen et al., 2006). The paradigm of quorum sensing states that AHLs are constitutively produced at low cell densities. The AHLs will then accumulate in the environment, in proportion to the increase in bacterial population. At a certain threshold concentration of AHL, the molecules will be able to bind to its respective receptor and a series of target gene regulations will be activated (Juhas et al., 2005). This system of regulation ensures that bacteria are able to form organised communities and to exchange information with other members to coordinate cellular activities. Among the processes regulated by quorum sensing are the synthesis of secondary metabolites, enzymes and virulence factors which allow bacteria to colonize various ecological niches (as reviewed by Juhas et al. 2005).
In P. aeruginosa, the quorum sensing systems are made up of the two AHL systems: las and rhl systems; and one non-AHL system: 2-heptyl-3-hydroxy-4-quinolone (PQS) (Pesci et al, 1999). The QS systems in P. aeruginosa regulate about 6-10% of the bacterial genome, indicating that this system play an important role in the survival of the bacteria (Arevalo-Ferro et al., 2003, Todar, 2004). QS in P. aeruginosa is important in pathogenicity as it ensures the bacteria do not express pathogenic traits until population has reached its critical density to be able to overwhelm host defences and establish an infection (Arevalo-Ferro et al., 2003). Among the virulence factors regulated by QS are proteases, exotoxin A, rhamnolipids, pyocyanin and sideophores (Wagner et al., 2003). In this report, the QS system is discussed in terms of its importance in the virulence of the organism as it causes serious implications in immunocompromised patients.
Mode of synthesis
The las system in P. aeruginosa quorum sensing comprise of LasI and LasR, where LasI synthesises the signal molecule N-(3-oxo-dodecanoyl)-L-homoserine lactone (3-oxo-C12- HSL) and LasR acts as a transcriptional regulator.
The second quorum sensing system, the rhl system, is comprised of RhlI and RhlR, where RhlI synthesises the production of signal molecule N-butanoyl-L-homoserine lactone (C4-HSL) and RhlR is the transcriptional regulator (Erikson et al., 2002, Rasmussen et al., 2006). LasR and RhlR are able to induce transcription of their own genes, creating a positive feedback loop that increases AHL production and dissemination (Schuster et al., 2006).
The PQS signalling system links the las and rhl systems. LasI and RhlI have been shown to control the synthesis of PQS which in turn, controls the expression of RhlI and RhlR. PQS gene transcription is positively regulated by LasR, and at the same time, it is negatively regulated by rhl. Therefore, the ratio of 3-oxo-C12-HSL:C4-HSL is important (as reviewed by Juhas et al., 2005). The las and rhl systems do not act independently; they are arranged in a hierarchical fashion where 3-oxo-C12-HSL positively regulates the rhl system (Erikson et al., 2002). Figure 1 represents a simplified depiction of the las and rhl system.

Figure 1 Quorum sensing in P. aeruginosa. The las act in a hierarchical fashion, whereby it controls the rhl. The LasR/3-oxo-C12-HSL complex activates the transcription of rhlR. Additionally, 3-oxo-C12-HSL can also block activation of RhlR by C4-HSL. Both quorum signaling systems regulate the expression of various genes (lasB: LasB elastase, lasA: LasA elastase, toxA: exotoxin A, aprA: alkaline protease, xcpP and xcpR: genes of the xcp secretory pathway, rhlAB: rhamnosyltransferase and rpoS: stationary phase sigma factor (Van Delden et al., 1998).
Structure
The structures of the AHL molecules and the PQS molecule are shown in Figure 2.
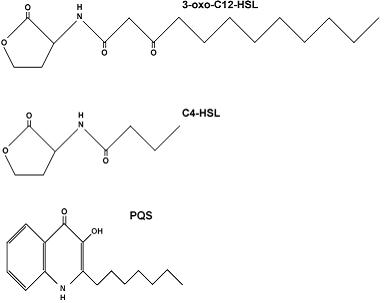
Fig. 2. Structures of PQS and AHLs exploited for cell-to-cell communication by P. aeruginosa (Juhas et al., 2005).
Biological Roles
The las system is mainly involved in the regulation of various virulence factors, while the rhl system regulates a broad spectrum of P. aeruginosa genes (Juhas et al., 2005). In CF patients, lasR transcripts have been detected in the sputum samples, and this accumulation correlated with lasA, lasB, toxA, arpA, lasI and rhlR mRNA transcripts, which are virulence factors (Pesci et al, 1997, Rumbaugh et al, 1999), indicating a functional link on the regulation of these genes. RhlR has been shown to bind to a specific upstream sequence of the rhlAB gene independently of the presence or absence of C4-HSL. When C4-HSL is present, transcription of rhlAB is activated, while transcription is repressed in the absence of C4-HSL (as reviewed by Juhas et al., 2005). C4-HSL has also been shown to regulate virulence genes such as lasB, rpoS, rhlA and rhlI (Rumbaugh et al., 1999). The PQS signalling system is able to exert antimicrobial activity and is shown to promote biofilm formation (as reviewed by Juhas et al., 2005).
Quorum sensing also play a part in host-pathogen interactions by modulating the host immune system. The AHL mainly responsible in immunomodulation of the immune system is 3-oxo-C12-HSL (Wagner et al., 2006). It is able to induce inflammation in infected hosts, which produces further damage. When 3-oxo-C12-HSL is produced, immune cells, such as lymphocytes, macrophages and antibodies are activated. The long chain AHL of 3-oxo-C12-HSL suppresses IL12 and TNFα secretion, skewing the T cell response to a Th2 type response (Telford et al., 1998). 3-oxo-C12-HSL has also been shown to induce Cox2 [an inflammation associated enzyme in macrophages]. Cox2 induces inflammation, fever and pain, which enables the pathogen to disseminate and cause septicemia. Therefore, this shows that 3-oxo-C12-HSL does not only regulate virulence genes, but it is a virulence factor by itself (Smith et al., 2002). 3-oxo-C12-HSL, which controls PQS production through the regulation of pqsH show that these compounds synergise to reduce T cell proliferation at sub-cytotoxic doses (Pritchard, 2006). PQS is able to act in the T cell signalling pathway by enhancing IL2 production, which in turn inhibits T cell proliferation. This suggests that the pathogen is capable of influencing the immune system to optimise its survival (Pritchard 2006, Smith et al., 2003).
To show that 3-oxo-C12-HSL and C4-HSL are directly accountable for pathogenicity, mutations in the las and rhl systems were made and the effects on pathogenicity were observed. Various experiments have shown that there is a general decrease in virulence in these mutants. In lung infection models, a lasI and rhlI double mutant produced less virulence factors, resulting in a faster immune response, stronger oxidative bursts of blood PMNs and accumulate antibodies faster (Rasmussen et al., 2006). In burn wound infections, lasI and rhlI double mutants showed reduced virulence as the pathogen was less efficient in dissemination compared to its wild type counterpart. This is because virulence factors, such as alkaline protease and elastase which interfere with phagocytosis of neutrophils are absent (Rumbaugh et al., 1999). In acute pulmonary infections (Pearson et al., 2000), burn wound infections (Rumbaugh et al., 1999) and chronic lung infections (Wu et al., 2001), mutants in QS genes have been able to reduce mortality compared to wild type infections. In a study done by Arevalo-Ferro et al. (2003), lasI and rhlI double mutant showed that cellular protein composition was significantly reduced compared to the cell’s transcriptome. This suggests that the LasR and RhlR transcriptional regulators are able to activate or repress the transcription of target genes. However, it should be noted that double mutants are not completely avirulent. Virulence of the bacteria is multi-factorial and quorum sensing is only a system that plays a part in virulence (Smith et al., 2003).
Practical Applications
The understanding of quorum sensing in P aeruginosa is important as it controls one third of the virulence genes. Hence, it can be exploited as a target for antimicrobials. Mutations made in the QS regulatory genes have been shown to reduce virulence and mortality. There are three ways to interfere with the QS system: blocking AHL synthesis, degrading the AHL signal molecule or inhibiting the binding of the signal molecule to the receptor (Juhas et al., 2005, Rasmussen et al., 2006). These methods minimise the selective pressure for resistant bacteria as it does not affect its growth (Juhas et al., 2005).
As precursors such as acyl-ACP and SAM are used in the synthesis of the AHL molecules, analogues of SAM, such as Holo ACP, L/D-S-adenosyl-homocystein, sinefugin and butyryl-SAM can inhibit synthesis of AHL synthetases, such as RhlI (as reviewed by Juhas et al., 2005). Therefore, there will be no accumulation of AHLs despite the increase in bacterial population. The AHL signal molecule could be degraded using chemical, enzymic or metabolic methods (Rasmussen et al., 2006). AiiA enzyme from Bacillus sp can inactivate AHLs by hydrolysing the lactone bond, also known as quorum quenching (as reviewed by Juhas et al., 2005).
Lactonolysis, or ring opening of the AHL lactones could be triggered by high pH as well as high temperatures. These result in lowered amounts of bioactive AHLs (Lee et al., 2002). Activation of the transcription regulators could be blocked using analogues of signalling molecules. Substitutions at the 3-oxo-C6-HSL acyl side chain are able to displace C6-HSL from activating the regulator (as reviewed by Juhas et al. 2005). The HSL ring can also be replaced with an alternative ring structure, such as exchanging the lactone ring for the amino cycloalcohol or amino cycloketone. These were shown to inhibit QS controlled expression of a plasI-gfp fusion and selected virulence factors (Smith et al., 2003).
Antimicrobials produced by other organisms could also block QS signalling. Delisea pulchra produces furanone, which is able to block cell-to-cell communication and is shown to be able to clear P. aeruginosa lung infections in mouse models (as reviewed by Juhas et al., 2005). QS inhibitors, such as para-benzoquinone and 4-nitro-pyridine-N-oxide (4-NPO) can also be used to target RhlR and LasR (Rasmussen et al., 2005). Eukaryotic organisms, such as fungi have also been shown to produce antimicrobial compounds such as patulin and penicillic acid to downregulate 45-60% of QS regulated genes.
QS systems are important in regulating the virulence of P. aeruginosa. Therefore, continued research to produce novel antibiotics that will be able to efficiently target this system would be greatly beneficial to reduce nosocomial infections and reduce mortality in immunocompromised patients.
References
Arevalo-Ferro, C., M. Hentzer, G. Reil, A. Görg, S. Kjelleberg, M. GIvskov, K. Riedel, and L. Eberl. 2003. Identification of quorum-sensing regulated proteins in the opportunistic pathogen Pseudomonas aeruginosa by proteomics. Environmental Microbiology. 5: 1350-1369.
Erikson, D. L., R. Endersby, A. Kirkham, K. Stuber, D. D. Vollman, H. R. Rabin, I. Mitchell, and D. G. Storey. 2002. Pseudomonas aeruginosa quorum-sensing systems may control virulence factor expression in the lungs of patients with cyctic fibrosis. Infection and Immunity. 70: 1783-1790.
Heurlier, K., V. Dénervaud, and D. Haas. 2006. Impact of quorum sensing on fitness of Pseudomonas aeruginosa. International Journal of Medical Microbiology. 296: 93-102.
Juhas, M., L. Eberl, and B. Tümmler. 2005. Quorum sensing: the power of cooperation in the world of Pseudomonas. Environmental Microbiology. 7: 459-471.
Lee, S. J., S.Y. Park, J.J. Lee, D.Y. Yum, B.T. Koo and J.K. Lee. 2002. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Applied Environmental Microbiology. 68: 3919–3924.
Pearson, J. P., M. Feldman, B. H. Iglewski, and A. Prince. 2000. Pseudomonas aeruginosa cell-to-cell signalling is required for virulence in a model of acute pulmonary infection. Infection and Immunity. 68: 4331-4334.
Pesci E. C., Pearson J. P., Seed P. C., Iglewski B. H. 1997. Regulation of las and rhl quorum sensing in Pseudomonas aeruginosa. Journal of Bacteriology 179:3127–3132
Pritchard, D. I. 2006. Immune modulation by Pseudomonsa aeruginosa quorum-sensing signal molecules. International Journal of Medical Microbiology. 296: 111-116.
Rasmussen, T. B., and M. Givskov. 2006. Quorum-sensing inhibitors as anti-pathogenic drugs. International Journal of Medical Microbiology. 296: 149-161.
Rasmussen, T. B., T. Bjarnsholt, M.E. Skindersoe, M. Hentzer, P. Kristoffersen, M. Kote, J. Nielsen, L. Eberl and M. Givskov. 2005. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. Journal of Bacteriology. 187: 1799–1814.
Rumbaugh, K. P., J. A. Griswold, B. H. Iglewski, and A. N. Hamood. 1999. Contribution of quorum sensing to the virulence of Pseudomonas aeruginosa in burn wound infections. Infection and Immunity. 67: 5854-5862
Schuster, M., and E. P. Greenberg. 2006. A network of networks: quorum sensing gene regulation in Pseudomonas aeruginosa. International Journal of Medical Microbiology. 296: 73-81.
Smith, R. J., S. G. Harris, R. Phipps, and B. Iglewski. 2002. The Pseudomonas aeruginosa quorum-sensing molecule N-(3-Oxododecanoyl)Homoserine Lactone contributes to virulence and induces inflammation in vivo. Journal of Bacteriology. 184: 1132-1139.
Smith, R. S., and B. H. Iglewski. 2003. P. aeruginosa quorum-sensing systems and virulence. Current Opinion in Microbiology. 6: 56-60.
Stover, C. K., X. Q. Pham, A. L. Erwin, A. L. Erwin, S. D. Mizoguchi, P. Warrener, M. J. Hickey, F.S. L. Brinkman, W. O. Hufnagle, D. J. Kowalik, M. Lagrou, R. L. Garber, L. Goltry, E. Tolentino, S. Westbrock-Wadman, Y. Yuan, L. L. Brody, S. N. Coulter, K. R. Folger, A. Kas, K. Larbig, R. Lim, K. Smith, D. Spencer, G. K.-S. Wong, Z. Wu, I. T. Paulsenk, J. Reizer, M. H. Saier, R. E. W. Hancock, S. Lory, and M. V. Olson. 2000. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature. 406: 959-964
Telford, G., D. Wheeler, P. Williams, P.T. Tomkins, P. Appleby, H. Sewell, G. Stewart, B.W. Bycroft and D.I. Pritchard, 1998. The Pseudomonas aeruginosa quorum-sensing signal molecule N-(3-oxododecanoyl)-L-homoserine lactone has immunomodulatory activity. Infection and Immunity. 66: 36–42.
Todar’s Online Textbook of Bacteriology. http://textbookofbacteriology.net/pseudomonas.html
Van Delden, C., and B. H. Iglewski.1998.Cell-to-cell signaling and Pseudomonas aeruginosa infections. Emerging Infectious Diseases 4: 551-560
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I. & Iglewski, B. H. 2003. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. Journal of Bacteriology. 185: 2080–2095.
Wagner, V. E., J. G. Frelinger, R. K. Barth, and B. H. Iglewski. 2006. Quorum sensing: dynamic response of Pseudomonas aeruginosa to external signals. Trends in Microbiology 14: 55-58
Wu, H., Z. Song, M. Givskov, G. Doring, D. Worlitzsch, K. Mathee, J. Rygaard, and N. Høiby. 2001. Pseudomonas aeruginosa mutations in lasI and rhlI quorum sensing systems result in milder chronic lung infection. Microbiology. 147: 1105-1113.
posted by GMO Pundit at 8:36 pm
![]()
![]()

1 Comments:
Ah that one
Here it is. After the talk I read it again, its really good
Science. 2006 Feb 24;311(5764):1113-6.
Bacterial small-molecule signaling pathways.
Camilli A, Bassler BL.
Bacteria use diverse small molecules for extra- and intracellular signaling.
They scan small-molecule mixtures to access information about both their extracellular environment and their intracellular physiological status, and based on this information, they continuously interpret their circumstances and react rapidly to changes. Bacteria must integrate extra- and intracellular signaling information to mount appropriate responses to changes in their environment. We review recent research into two fundamental bacterial small-molecule signaling pathways: extracellular quorum-sensing signaling and intracellular cyclic dinucleotide signaling. We suggest how these two pathways may converge to control complex processes including multicellularity, biofilm formation, and virulence. We also outline new questions that have arisen from recent studies in these fields.
Post a Comment
<< Home